Статистическим ансамблем физической системы называется набор всевозможных состояний данной системы, отвечающих определённым критериям. Примерами статистического ансамбля является большой канонический ансамбль, описывающий состояния системы с переменным числом частиц. Большой канонический ансамбль соответствует физической системе, которая обменивается энергией и частицами с окружающей средой, но находится с ней в тепловом равновесии. Если термодинамическая система может обмениваться со средой частицами, а не только энергией, то со временем устанавливается не только тепловое равновесие между системой и средой, но и равновесие по составу.
CDMA (Code Division Multiple Access — множественный доступ с кодовым разделением (МДКР)) — технология связи, обычно радиосвязи, при которой каналы передачи имеют общую полосу частот, но разные кодирующие последовательности. Наибольшую известность на бытовом уровне получила после появления сетей сотовой мобильной связи, её использующих.
При кодовой модуляции применяется техника расширения спектра с множественным доступом. Она позволяет увеличить пропускную способность при неизменной мощности сигнала. Передаваемые данные комбинируются с более быстрым шумоподобным псевдослучайным сигналом с использованием операции побитового взаимоисключающего ИЛИ
Расширение спектра — способ повышения эффективности передачи информации с помощью модулированных сигналов через канал с сильными линейными искажениями (замираниями), приводящий к увеличению базы сигнала в частотной области.
В WI-FI, например, применяется расширение спектра методом прямой последовательности (Direct sequence spread spectrum, DSSS). Метод по эффективности превосходит псевдо случайную перестройку рабочей частоты, но сложнее в реализации. Суть метода заключается в повышении тактовой частоты модуляции, при этом каждому символу передаваемого сообщения ставится в соответствие некоторая достаточно длинная псевдослучайная последовательность (ПСП). Метод используется в таких системах как CDMA и системах стандарта IEEE 802.11.
Для выделения сигнала из шумов и помех применяются различные методы и технические приемы. Среди них наибольшее распространение получили корреляционный прием, оптимальная фильтрация и накопление сигналов.
Метод корреляционного приема сигналов. Корреляционный прием основан на сравнении поступающего сигнала с набором сигналов-образцов. При этом показателем совпадения является значение корреляционной функции на периоде следования сигнала (в течение такта передачи).
Как следует из теории оптимальных методов радиоприема, корреляционный приемник обеспечивает наивысшую вость, т.е. максимально возможное отношение сигнала к шуму на выходе приемника. Единственным недостатком корреляционного приемника является сложность практической реализации.

Структурная схема корреляционного приемника
Структура радиоинтерфейса сети 5G-NR
В 5G-NR используются OFDM поднесущие с различной шириной спектра – 15кГц, 30кГц, 60кГц, 120кГц и 240кГц – см. Табл. (в отличии от сетей LTE, где используется стандартная ширина спектра поднесущей, равная 15кГц). Использование различных нумерологий открывает широкие возможности для гибкой настройки поведения сети при предоставлении тех или иных услуг, например, для приложений критичных к уровню задержек (URLCC), уместно использовать поднесущие с широким спектром при меньшей длительности символа, и наоборот, при передаче трафика широкополосного доступа в Интернет и низкоскоростного трафика Интернета вещей – использовать «узкие» поднесущие.
| u (нумерология) | ∆F=2u∙15 [кГц] | Циклический префикс |
| 0 | 15 (соответствует LTE) | Normal |
| 1 | 30 | Normal |
| 2 | 60 | Normal, Extended |
| 3 | 120 | Normal |
| 4 | 240 | Normal |
В 5G-NR кол-во OFDM символов, приходящихся на один слот
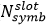
не зависит от нумерологии и, по аналогии с сетями 4G-LTE, определяется только типом циклического префикса:
- 14 символов для нормального (Normal)
- 12 символов для расширенного (Extended) префикса.

Для наглядности ,приведена диаграмма OFDM-символов на сетке координат «частота»-«время»

Каждый OFDM символ слота может иметь тип «downlink» или «D» (нисходящий канал), «uplink» или «U» (восходящий канал) и «flexible» или «X» (гибкая конфигурация). В Табл. 3 приведены доступные форматы распределения OFDM символов между восходящим и нисходящим направлениями для нормального циклического префикса (Таблица 11.1.1-1 3GPP TS 38.213). Процедура конфигурации слотов описана в спецификации 3GPP TS 38.213
По аналогии с 4G-LTE единицей частотно-временного ресурса в сетях 5G-NR является ресурсный блок – RB (Resource Block). Каждому абонентскому терминалу на определенный период времени для приема/передачи данных выделяется некоторое, определенное планировщиком системы, кол-во ресурсных блоков. Каждый ресурсный блок в частотной области содержит

смежных поднесущих частот. Ширина полосы частот одного ресурсного блока зависит от используемой нумерологии
| u (нумерология) | Поднесущая, ∆F [кГц] | 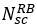 | Ресурсный блок [кГц] |
| 0 | 15 | 12 | 180 |
| 1 | 30 | 12 | 360 |
| 2 | 60 | 12 | 720 |
| 3 | 120 | 12 | 1 440 |
| 4 | 240 | 12 | 2 880 |
Основой системы радиодоступа сети 5G-NR является технология мультиплексирования с ортогональным частотным разделением каналов OFDM (Orthogonal Frequency Division Multiplexing).
В нисходящем направлении от базовой станции к абонентскому терминалу (Downlink – DL) используется простой метод CP-OFDM (Cyclic Prefix OFDM), при котором каждый пользовательский канал на определенном временном интервале состоит из совокупности параллельно излучаемых сигналов на множестве поднесущих частот.
В восходящем направлении от абонентского терминала к базовой станции (Uplink – UL) возможны два варианта – CP-OFDM (по аналогии с DL) и DFT-s-OFDM (схожий с технологией SC-FDMA сети 4G-LTE). Последний включает блок «Transform Precoding», фактически реализующий операцию дискретного преобразования Фурье (DFT) и позволяет снизить пик фактор (отношение максимального и среднего уровней мощности).

Возможные схемы модуляции в 5G-NR – π/2-BPSK, QPSK, 16QAM, 64QAM, 256QAM. При этом модуляция π/2-BPSK (неиспользуемая в сетях 4G-LTE) доступна только в восходящем (UL) канале в режиме DFT-s-OFDM и актуальна для энергоэффективных сетей интернета вещей.
Пространственные потоки (MIMO) 5G
Одним из ключевых нововведений в технологии 5G является использование массивных многоантенных систем (Massive Multiple-Input-Multiple-Output) под которыми принято понимать системы, имеющие более 8-ми управляемых антенн и приемо-передающих трактов. Использование Massive MIMO позволяет с одной стороны увеличить емкость сети (в 5G-NR доступное кол-во параллельных пространственных потоков данных в нисходящем канале «DL» – 8; в восходящем «UL» – 4), с другой – повысить помехоустойчивость за счет использования техники формирования направленных лучей (beamforming).
Главное в сказанном. Каждая Massive Multiple-Input-Multiple-Output обеспечивает формирование направленной диаграммы антенны на объект, с которым устанавливается связь, а три соседних вышки с антеннами Multiple-Input-Multiple-Output обеспечивают точное обнаружение местоположения объекта методом триангуляции.

С учетом длины волны — точность засечки фигуры пеленгации для подвижного объекта связи в существующей полосе ВРК-15 составит 0,5 — 1 сантиметр, в среднесрочной перспективе — доли миллиметра.

Рассмотрим пару любопытных документов


Эволюция микрочипов ДНК
Микрочип ДНК (также широко известный как ДНК-чип или биочип) представляет собой набор микроскопических пятен ДНК, прикрепленных к твердой поверхности. Ученые используют микрочипы ДНК для измерения уровней экспрессии большого количества генов одновременно или для генотипирования нескольких областей генома.
Каждое пятно ДНК содержит пикомолы (10-12молей) определенной последовательности ДНК, известные как зонды (или репортеры или олигосы). Это может быть короткий участок гена или другого элемента ДНК, который используется для гибридизации образца кДНК или кРНК (также называемой антисмысловой РНК) (называемой мишенью) в условиях высокой жесткости.
Гибридизацию зонд-мишень обычно обнаруживают и количественно оценивают путем обнаружения меченых флуорофором, серебром или хемилюминесценцией мишеней для определения относительного обилия последовательностей нуклеиновых кислот в мишени.
Первоначальные массивы нуклеиновых кислот представляли собой макро-массивы размером примерно 9 × 12 см, а первый компьютерный анализ на основе изображений был опубликован в 1981 году.
Эта же технология может использоваться в качестве SNPs-массивов для анализа полиморфизмов при сердечно-сосудистых заболеваниях, раке, патогенах и GWAS-анализе и для идентификации структурных вариаций и измерения экспрессии генов.
Где берут образцы для таких анализов — написано в отчете Европейской комиссии.
Из этого досье следует, что мы с Вами едим ГМО свеклу H7 от Monsanto в переработанном на сахар виде.
Этот инструмент помогает контрольным лабораториям в разработке стратегий скрининга и интерпретации результатов с помощью набора из трех интерфейсов:
ГМО-матрица Этот интерфейс может строить двумерные матрицы (выбранные события ГМО против выбранных методов ГМО) для визуализации аналитического покрытия и выявления потенциальных пробелов в подходе скрининга пользователей.
GMO-Event finder Этот интерфейс позволяет идентифицировать потенциальные ГМО, присутствующие в образце, на основе набора методов обнаружения, определенных пользователем, и полученных положительных и отрицательных экспериментальных результатов.
Пластины Этот интерфейс предоставляет веб-инструмент для интерпретации результатов готового к использованию многоцелевого скринингового устройства, разработанного JRC и известного как pre-spotted plate (PSP). Это устройство позволяет проводить тестирование на наличие нескольких ГМО-мишеней в одном ПЦР-эксперименте.
Основным принципом микрочипов является гибридизация между двумя нитями ДНК, свойство комплементарных последовательностей нуклеиновых кислот специфически соединяться друг с другом путем образования водородных связей между комплементарными парами нуклеотидных оснований. Большое количество комплементарных пар оснований в нуклеотидной последовательности означает более плотную нековалентную связь. соединение между двумя нитями.
После смывания неспецифических связующих последовательностей гибридизованными останутся только сильно спаренные нити. Флуоресцентно меченые целевые последовательности, которые связываются с зондовой последовательностью, генерируют сигнал, который зависит от условий гибридизации (таких как температура) и промывки после гибридизации. Общая сила сигнала от пятна (признака) зависит от количества связывания целевого образца с зондами, присутствующими в этом месте. Микрочипы используют относительную количественную оценку, при которой интенсивность признака сравнивается с интенсивностью того же признака при других условиях, а идентичность признака определяется его положением.
Для получения дополнительной информации следует почитать:
- «Разработка и применение готовой к использованию системы ПЦР для скрининга ГМО».
Пищевая химия, 2016 15 июня; 110-119. ссылка - «JRC GMO-Matrix: веб-приложение для поддержки стратегий обнаружения генетически модифицированных организмов.»
BMC Bioinformatics. 2014 Dec 30;15(1):6592. ссылка
Микрочипы ДНК можно использовать для обнаружения ДНК (как при сравнительной геномной гибридизации
или обнаружения РНК (чаще всего в виде кДНК после обратной транскрипции), которая может быть переведена или не переведена в белки. Процесс измерения экспрессии генов с помощью кДНК называется анализом экспрессии или профилированием экспрессии.
Правовая основа для ГМО

Объединенный исследовательский центр (ОИЦ) Европейской комиссии был назначен Референтной лабораторией Европейского Союза по генетически модифицированным пищевым продуктам и кормам (EURL GMFF) (1) в соответствии с Постановлением (ЕС) № 1829/2003.
В соответствии с его положениями СИЦ оказывает помощь консорциум Национальных референс-лабораторий (НРЛ), именуемый “Европейской сетью лабораторий по ГМО” (В соответствии с Регламентом (ЕС) № 882/2004 ОИЦ также обозначается как EURL GMFF (2) для официального контроля, осуществляемого с целью обеспечения проверки соблюдения законодательства о кормах и пищевых продуктах, правил охраны здоровья животных и благополучия животных.
Этот Регламент был отменен Регламентом (ЕС) 2017/625 об официальном контроле и других официальных мероприятиях, осуществляемых для обеспечения дополнительной проверки правил охраны здоровья растений и средств защиты растений.
Ниже приведен список правовых актов ЕС по ГМО имеющих отношение к задачам и деятельности EURL GMFF:
- Регламент (ЕС) № 1829/2003 о ГМ-продуктах питания и кормах официально устанавливает EURL и определяет ее задачи
- Регламент (ЕС) 2017/625на официальных мер контроля и другими официальными мероприятиями, отменяющий регламент (EC) № 882/2004 на официальном контроля, обеспечивает проверку соответствия кормов и пищевого законодательства, ветеринарно-санитарных, животных и здоровья растений и защиты растений, правил и определены задачи на лес отслеживания для гармонизации ГМО-контроль деятельности на протяжении государств-членов ЕС.
- Комиссия по имплементации Регламента (ЕС) № 120/2014 о внесении поправок в Регламент (ЕС) № 1981/2006 обновляет список назначенных Национальных референтных лабораторий, поддерживающих EURL GMFF для валидации метода.
- Регламент (ЕС) № 619/2011, устанавливающий методы отбора проб и анализа для официального контроля кормов в отношении наличия генетически модифицированного материала, в отношении которого находится на рассмотрении процедура авторизации или срок действия разрешения на который истек, делает валидированный метод обнаружения предварительным условием для низкого уровня присутствия в кормах других разрешенных ГМО.
Другие соответствующие правовые акты
- Директива 2001/18 о преднамеренном выбросе в окружающую среду генетически модифицированных организмов и отмене Директивы Совета 90/220/EEC.
Чрезвычайные ситуации в Европейской комиссии тоже бывают Emergencies / Unauthorised
- Page dedicated to GM wheat
- Verification Report on the extraction and analysis of GM pollen DNA in honey
- Page dedicated to Commission implementing decision 2013/287/EU (emegency measures regarding unauthorised GM rice in rice products originating from China)
- Page dedicated to CDC Triffid flax (FP967)
- Page dedicated to Bt63 rice
- Page dedicated to E32 maize
- Page dedicated to LLRICE601 rice
- Page dedicated to BT10 maize
- Page dedicated to Oilseed Rape Oxy-235
Введение в microarrays
ДНК-микрочипы могут быть использованы для обнаружения ДНК (как при сравнительной геномной гибридизации) или РНК (чаще всего в виде кДНК после обратной транскрипции), которая может быть переведена или не переведена в белки. Процесс измерения экспрессии генов с помощью кДНК называется анализом экспрессии или профилированием экспрессии.
Значительными участниками глобального рынка анализа экспрессии генов являются
General Eletric Company, Illumina, Inc., Bio-Rad Laboratories, Inc., F. Hoffmann-La Roche Ltd, Thermo Fisher Scientific, Pacific Biosciences of California, Inc., Oxford Gene Technology, BGI, PerkinElmer Inc., Agilent Technologies, Inc., QIAGEN, Eurofins Scientific, Genotypic Technology Pvt Ltd, Danaher, Macrogen Inc.
Эволюция microarrays к nanoarrays
Размеры подложки nanoarrays не превышают размеров осаждаемых на них молекул

В самых общих чертах развитие технологии микрочипов шло по трем направлениям: увеличение числа тестов (биомолекул или клеток) на одном и том же чипе;
- увеличение числа тестируемых биомолекул на одной и той же единице площади (то есть плотности);
- увеличение сложности биочипов, причем предлагалось множество альтернативных конструкций.
Первые две тенденции идут по ступеням эволюции собственно микрочипов, то есть более крупных чипов и более высокой плотности на чипе, но последнее сходство (то есть «умная» конструкция) должно быть проанализировано более подробно. В то время как полупроводниковая технология очень рано ввела в свою историю «чемпионское» устройство, то есть биполярный и более поздний КМОП-транзистор, технология микрочипов еще не имеет » чемпиона’.
Из этого следует, что технология микрочипов еще не достигла своей зрелости, со всеми преимуществами (например, эмерджентные инновации) и недостатками (например, ди-культовая стандартизация), которые возникают на этой все еще развивающейся стадии.
Однако, если смотреть с другой стороны, технология микропреломления гораздо ближе к «технологическому кризису». На протяжении десятилетий утверждалось, и каждый раз доказывалось, что полупроводниковая технология остановится из-за неспособности литографии печатать меньшие объекты в темпе, заданном неумолимым законом Мура, т. е.
Вдвое уменьшать критический размер печати на чипе каждые 18 месяцев. Помимо кажущейся бесконечной способности микролитографии раздвигать пределы разрешающей способности, фундаментально кризис всегда был далеко.
Микроэлектроника и – в настоящее время – наноэлектроника «работают» с электронами (которые намного меньше 1 нм), в то время как самая продвинутая литография призвана печатать особенности многих десятков нанометров.
Даже если мы рассмотрим квантовые эффекты, нынешняя литография может печатать объекты, которые по меньшей мере в десять раз превышают критический технологический барьер. С другой стороны, отдельные ДНК и белки, мельчайшие «строительные элементы» микрополосков, имеют размеры от нескольких до нескольких десятков нанометров.
Следовательно, продвинутая литография уже способна печатать объекты, которые находятся на том же порядке величины, если не меньше, чем «модули», подлежащие печати. Кроме того, для массивов клеток и тканей разрешение паттернов не является проблемой.
Во-вторых, способность обращаться к единичным биомолекулам позволяет концептуализировать совершенно разные микро-и наноматериалы.
Например, вместо «пассивных», «одноразовых» массивов, где биомолекулы исследуются (подобно функции «чтения») один раз, можно подумать об «активных», «многократных» массивах, где биомолекулы выполняют повторяющиеся функции, например «вычисления».
Другой уровень сложности может быть добавлен, если мы перейдем от «статических» массивов, где биомолекулы находятся и зондируются в одном месте, к «динамическим» массивам с биомолекулами, которые перемещаются либо вбок либо кругом, чтобы выполнять повторяющиеся задачи, например, зондирование, выработку энергии и снова вычисление.
В принципе, существуют две основные стратегии изготовления наночастиц, которые являются общими для микро/нанолитографии как для микрочипов, так и для микрочипов.
- Во-первых, можно изменять свойства области, например, с помощью света, создавая различные химические функции или гидрофобности локально. Впоследствии этот «остров» используется для дальнейшего изготовления, например, иммобилизации целевой биомолекулы. Эта стратегия изготовления имеет определенные фундаментальные ограничения с точки зрения достижимого разрешения.
- Во–вторых, можно локально осаждать химические виды (например, целевые биомолекулы в растворе) механическим способом непосредственно на поверхности, например, наноразмерным «карандашом» – стратегия, аналогичная нескольким «литографиям нового поколения».
Осаждение механическим способом также может быть выполнено бесконтактным способом с использованием технологий, разработанных на основе технологии струйного принтера.
Современные методы получения микрочипов варьируются в зависимости от конкретного применения и включают контактные и бесконтактные методы выделения олигонуклеотидов или кДНК, а также комбинацию фотолитографии и синтеза олигонуклеотидов in situ.
Однако без значительных инвестиций в высококачественные проекционные системы традиционные методы литографии не могут создавать объекты в диапазоне 150-200 нм из-за дифракции света. Экстремальная УФ–литография и другие стратегии фотолитографии следующего поколения могут обеспечить требуемое разрешение, но при постоянно возрастающих затратах на маски и средства изготовления, а также при работе в все более жестких условиях, которые могут быть несовместимы с биологическими материалами. В частности, в конечном итоге станет экономически невыгодным уменьшать размеры пятен микрочипов с помощью обычной фотолитографии.
Изготовление наночастиц с разрешением менее 100 нм
Комбинаторные nanoarrays, изготовленные методом микроабляции
Биомолекулы, в частности белки, сильно взаимодействуют с поверхностями, на которых они иммобилизованы. Наноструктуры обладали бы как способностью исследовать большие биомолекулы по отдельности, поскольку они имеют сопоставимые размеры с исследуемыми биомолекулами, так и способностью делать это зондирование в значительной степени параллельным, поскольку наноструктуры поддаются большой плотности площади.
В целом микрообработка не способна производить наноструктуры, но в последнее время лазерная микроабляция используется для изготовления структур, которые имеют микронные размеры в поперечном направлении, но наноразмерные в вертикальном.
Микро–лунки изготавливаются путем локализованной лазерной абляции тонкого (десятки нм) слоя металла (например, золота), нанесенного на прозрачную полимерную (например, ПММА) пленку.
Микроабляция золота вызывает локальные химические и физические изменения в верхней поверхности полимера, а также более высокую удельную поверхность, которые взаимодействуют для достижения более высокой и воспроизводимой поверхностной концентрации белков в микро–лунках.
Метод изготовления может использовать последовательность локальной абляции и «заливки» белковым раствором или абляцию всего микроанализа с последующим «пространственно–адресным» осаждением различных белковых растворов пико-литровой пипеткой. Было замечено, что микроанализы, содержащие линейчатые микроструктуры, обеспечивают более высокую воспроизводимость и возможность кодирования информации (напр. тип белка, концентрация) через комбинацию вертикальных линий в режиме «штрих-код», «информационно-адресуемый», а не в пространственно-адресуемом режиме, как в классических микрочипах. Установлено, что «комбинаторная» природа внутренней поверхности каналов позволяет увеличить адсорбцию молекулярно различных белков от 3 до 10 раз по сравнению с адсорбцией на аналогичных плоских поверхностях при более высокой амплификации более мелких глобулярных белков.
Dip Pen NanolithographyTM
Существует метод прямой записи нанопаттернирования на основе SPM – «Dip Pen NanolithographyTM» (DPNTM) Основанный на обычной АСМ, ДПНТМ сочетает в себе разрешения, сравнимые и в некоторых случаях превосходящие разрешения электронно-лучевой литографии (ширина линии 15 нм), с широкой химической совместимостью, полученной при работе в условиях окружающей среды.
В типичном ДПНТМ в эксперименте обычный наконечник АСМ-зонда покрывают молекулой или «чернилами», которые рисуют узором, погружая наконечник в раствор этих молекул. При контакте наконечника с поверхностью молекулы осаждаются через водяной мениск, который конденсируется при контакте наконечника с подложкой. При таком диффузионном процессе более длительное время пребывания наконечника-подложки приводит к увеличению площади пятен рисунка.
Благодаря своей природе прямого осаждения ДПНТМ было показано, что процесс является очень общим как в отношении молекул, которые могут быть перенесены с наконечника АСМ на поверхность (небольшие органические поверхностно–активные вещества, заряженные макромолекулы, такие как сопряженные полимеры и белки, золь-гель образующие материалы и даже наночастицы), так и в отношении подложки (металлы, например золото; изоляторы, например оксид кремния; и полупроводники, например GaAs).
Основное требование для переноса заключается в том, что существует некоторое взаимодействие, ковалентное или физическое между чернилами и поверхностью. Например, алкантиолы образуют координационную связь с золотой поверхности.
Хотя существуют методы, которые могут быть использованы для получения чрезвычайно тонких структур на поверхности (например, электронно–лучевая или сфокусированная ионно-лучевая литография), проблема изготовления наноматериалов заключается в создании сложных узоров, состоящих из различных материалов, расположенных в точном положении относительно друг друга.
С помощью DPNTM можно использовать возможность записи и чтения химических паттернов высокого разрешения с помощью одного и того же инструмента. Таким образом, с помощью DPN TM можно генерировать множество химических или биологических паттернов с точной (5 нм) регистрацией выравнивания.
Среди методов нанесения рисунка, которые могут работать на субмикронных и суб–100 нм размерах, таких как электронная лучевая литография или методы контактной штамповки, DPNTM это единственная технология, которая может непосредственно осаждать биологические молекулы в условиях окружающей среды со сверхвысокой точностью и регистрацией.
Кроме того, эти молекулы могут осаждаться в окружающей или инертной среде без воздействия на них ионизирующего УФ–или электронно-лучевого излучения.
Кроме того, можно осаждать несколько различных типов молекул, не подвергая их воздействию жестких растворителей или химических травителей и не рискуя перекрестным загрязнением. Искомая химия осуществляется точно и только там, где она желательна.
Вычислительные наночастицы
Функция » классических’ био–массивов, будь то микро– или нано– массивы, заключается в предоставлении информации о биомолекулярном распознавании посредством стыковки зондирующих биомолекул с целевыми биомолекулами (или клетками), пространственно закодированными на поверхности массива. Но молекулярное распознавание может быть только первым из множества этапов, представляющих собой процесс вычисления, и в этом случае последняя конфигурация микрочипа представляет собой «решение». Хотя эти функциональные массивы не являются наноматериалами в смысле поперечного или вертикального разрешения, как описано в предыдущем разделе, они действительно выполняют свою функцию на наноуровне.
ДНК-вычисления-это новый метод физических вычислений, основанный на молекулярном распознавании комплементарных биомолекул (ДНК) и массивном параллелизме, который может быть достигнут с помощью циклов синтеза ДНК, ПЦР, лигирования, электрофореза и использования ограничительных ферментов. Этот новый вычислительный метод, по-видимому, особенно подходит для задач, которые не могут быть решены даже самыми передовыми традиционными электронными компьютерами, работающими последовательно. Традиционно их называют NP-проблемами, имея в виду экспоненциальное недетерминированное полиномиальное время, необходимое для достижения решения при линейном увеличении размера задачи.
Адлеман был первым, кто описал основанный на ДНК метод, который решает проблему пути Гамильтона (например, находит маршрут авиакомпании, проходящий через несколько городов и оптимально посещающий каждый только один раз) за полиномиальное время.
Компромисс в эксперименте Адлемана заключался в использовании большого числа «компьютеров» (то есть молекул ДНК), которые выполняют операции в массовом порядке параллельно времени (количество и тип физических процедурных шагов).
Узлы и пара между узлами были закодированы в нитях ДНК, которые самособираются во всех возможных сочетаниях после комплементарности Уотсона–Крика.
Некоторые из этих dsDNA могут содержать возможные решения, которые могут быть выбраны, усилены и обнаружены с помощью классических методов молекулярной биологии.
После первоначального доказательства концепции, проведенного Адлеманом, ДНК-вычислениям было уделено большое внимание из-за их потенциала для решения проблем, емкости хранения данных, энергоэффективных вычислений и нового математического взгляда на вычисления.
По существу, основными операциями вычисления ДНК являются:
- Амплификация;
- Слияние;
- Обнаружение;
- Последовательность-отдельная;
- Длина-отдельная;
- Положение-отдельная
Используя этот базовый математический аппарат, было предложено множество алгоритмов для решения конкретных задач с использованием ДНК-вычислений, среди которых решены:
- проблема выполнимости;
- проблема максимального отклика;
- проблема раскраски графа;
- взлом схемы шифрования данных;
- проблема коммивояжера;
- решение связности графа;
- проблема «рюкзака»
Решающим фактором, от которого зависит успех ДНК– вычислений в фазе растворения, является способность достигать очень малых частот ошибок для различных биохимических операций. Поскольку главная идея ДНК-вычислений заключается в выполнении массивно-параллельных операций, из этого следует, что эффективное вычисление опирается на максимально полный поиск пространства возможных решений (нитей ДНК).
Классически это достигается высоким соотношением доступных нитей ДНК на число возможных растворов.
По мере увеличения сложности задачи (выраженной в измерениях входных данных) это отношение уменьшается для данного начального количества ДНК, то есть среднее число нитей, кодирующих одно потенциальное решение, становится меньше.
Это накладывает требования на максимально допустимую частоту ошибок или, что эквивалентно, на минимальное количество необходимой ДНК. Таким образом, ДНК-вычисления, как они определены, страдают от проблемы «масштабируемости».
Это побудило искать средства для лучшего контроля частоты ошибок в операциях вычисления ДНК (например, ПЦР, гибридизация). Одним из путей улучшения экспериментального контроля во время экспериментов по вычислению ДНК является иммобилизация нитей ДНК на поверхности перед манипуляцией.
Технология микрочипов помогла перенести концепцию манипулирования молекулами ДНК для ДНК-вычислений с процессов, основанных на растворах, на процессы, основанные на поверхности.
Комбинаторные библиотеки ssDNA распознаются гибридизацией к их дополнениям (операция «марка»), что делает эти части двухцепочечными. Фермент (например, экзонуклеаза) разрушает не гибридизованные олигонуклеотиды (функция «разрушения»).
Наконец, ранее гибридизованные олигонуклеотиды регенерируются (операция «unmark»).
Все нити, которые не представляют решение, удаляются путем повторения операций «пометить», «уничтожить» и «снять», оставляя только «решение», связанное на поверхности. Наконец, раствор считывают через последовательность отсоединения от поверхности, ПЦР и дальнейшую гибридизацию с разработанным микрочипом. Frutos et al развили этот метод дальше, предложив использовать реакции ферментативного лигирования ДНК — » слов’ на поверхностях для вычисления ДНК.
Этот метод вычисления был использован Liu et al. для решения простого случая задачи 3–SAT, которая считается самой сложной из всех NP-задач. Решение задачи 3–SAT должно удовлетворять набору логических предложений, каждое из которых состоит из трех истинных или ложных переменных, связанных
с помощью логических операторов » или’. Эта проблема была решена в разумные сроки путем кодирования переменных в двоичных строках, которые, в свою очередь, были закодированы в строках ssDNA. Для n переменных существует 2n уникальных цепей «ответа’ (или «Ватсона»), например TGCGG = 001, дополненных уникальными цепями » Крика’. Решение принимается, если оно удовлетворяет всем логическим положениям формулы 3–SAT. Если нити ssDNA, представляющие все возможные растворы, иммобилизовать на поверхности золота, то добавление нитей Крика создаст комбинацию ss– и dsDNA. Нерастворенные ssDNA, которые не удовлетворяют первому предложению, закодированному в добавленных нитях Крика, разрушаются ферментами, оставляя все еще возможные решения, запертые в нитях dsDNA, которые впоследствии расплавляются, и процесс начинается снова для следующего предложения.
Последняя оставшаяся нить — это решение, которое декодируется в формате микрочипа.
Кроме синтеза нитей ДНК, вычисление происходит в 3k + 1 шагах для исследования всех возможностей 2n (k-число предложений). Эта процедура намного эффективнее, чем лучший обычный компьютерный алгоритм, который масштабируется как 1,33n (n = число переменных).
Чтобы представить ситуацию в перспективе, задача 3–SAT с 30 предложениями и 50 переменными была бы решена классически примерно за 1,6 миллиона шагов, но существуют методы, которые решают эту задачу за 91 шаг.
С математической точки зрения поверхностные вычисления ДНК-это конкурент решающим вычислениям ДНК.
Кроме того, количество операций в секунду ограничено более медленной кинетикой фермента и более низкой эффективностью гибридизации. Наконец, поверхностный метод не устраняет проблем масштабирования, поскольку различение одноосновных несоответствий становится более трудным по мере увеличения длины нити и операции не являются безошибочными. Наиболее серьезным из этих ограничений является потеря плотности информации. Необходимо либо увеличить площадь поверхности (напр. с помощью микробов вместо плоской поверхности) или попытаться использовать локальную трехмерную химию поверхности.
Динамические наночастицы
Другой характерной чертой » классических’ микро-и наночастиц является их одноразовое использование. Как только их функция, будь то простое молекулярное распознавание или биомолекулярное вычисление, выполняется и информация передается далее в соответствующие системы обработки информации, продукт –микрочип– устаревает, и поэтому микро/наночипы по существу являются одноразовыми устройствами (за заметным исключением подхода Nanogen, полученного из биосенсоров). Более продвинутые устройства будут разработаны для использования молекулярного распознавания , а не для, их функция, которая тогда была бы непрерывной, а не одно– о. Эти будущие устройства, которые будут работать в очень параллельном расположении, возможно, в формате микрочипа, будут содержать движущиеся элементы, приводимые в движение внешними средствами или, предпочтительно, самоходные. Первый вариант, то есть динамические устройства с внешним питанием, был запущен с помощью микрофлюидики и манипулирования магнитными шариками. Однако именно самоходные динамические устройства дают самые высокие ожидания технологических революций. К счастью, Природа предлагает несколько работающих моделей молекулярных двигателей, многие из которых испытаны на примитивных гибридных динамических наноустройствах.
Белковые молекулярные двигатели, работающие либо в паре, то есть линейные двигатели, либо в паре, то есть вращательные двигатели, преобразуют химическую энергию посредством гидролиза аденозинтрифосфата (АТФ) в механическую энергию или движение. Молекулярные двигатели, которые являются вездесущими белками, отвечают за такие разнообразные биологические функции, как движение и деление клеток, транспортировка везикул и мышечная функция.
Два экспериментальных метода, анализ подвижности и визуализация одной молекулы, манипуляция и измерение, привели к важным достижениям в понимании и количественной оценке функций молекулярных двигателей. Анализ подвижности, впервые проведенный около 15 лет назад, по сути, представляет собой примитивные наноустройства, работающие в «распределенном» формате микродвигателей, которые позволяют зондировать функции молекулярных двигателей «черным ящиком». С другой стороны, одномолекулярные методы позволяют измерять фундаментальные параметры, например — силы, и поэтому полезны для проектирования будущих наноустройств на основе молекулярных двигателей.
Роторные двигатели
Белковые молекулярные двигатели выполняют свою функцию посредством вращательного или линейного движения. Хотя было показано, что актиновые нити также совершают вращательное движение вдоль своей оси при скольжении по миозиновым функционализированным поверхностям, есть два двигателя, которые работают в истинно вращательном режиме, т. е. двигатель бактериального жгутика и фермент АТФ-синтазы/
Последний, по — видимому, является самым маленьким (приблизительно 12 нм), самым эффективным (генерирующим около 100 пН нм с почти 100% эффективностью) и самым быстрым (скорость вращения без нагрузки около 17 об / с) роторным двигателем. Все эти преимущества делают эту систему весьма привлекательной для использования в гибридных наноустройствах. АТФ-синтаза-это крупный фермент, который синтезирует АТФ в митохондриях. Подобные ферменты можно найти и в других организмах, например в хлоропластах растений и клеточных мембранах бактерий, причем последние особенно подходят для создания надежных гибридных наноустройств.
Литература, опубликованная до 2003 года.
- 1. Dammel, R. Diazonaphthoquinone–based Resists; SPIE Tutorial Texts; SPIE: Bellingham,1993; Vol. 11, Chapter 1
- 2. Pirrung, M. C. Angew. Chem. Int. Ed. 2002, 41, 1276–1289 116 Dan V. Nicolau et al.
- 3. Lander, E. S. Nature Genet. 1999, 21, 3–4
- 4. Xia, Y.; Rogers, J. A.; Paul, K. E.; Whitesides, G. M. Chem. Rev. 1999, 99, 1823–1848
- 5. Geppert, L. IEEE Spectrum 1996, 33–38
- 6. Ivanova, E., Wright, J.P., Pham, D.K., Filipponi, L., Viezolli, A., Nicolau, D.V. Langmuir, 18, 9539–9546, 2002
- 7. Liu, G.-Y.; Xu, S.; Qian, Y. Acc. Chem. Res. 2000, 33, 457–466
- 8. Xu, S.; Liu, G. Langmuir 1997, 13, 127–129
- 9. Shoer, J. K.; Crooks, R. M. Langmuir 1997, 13
- 10. Bruckbauer, A.; Ying, L.; Rothery, A. M.; Zhou, D.; Shevchuk, A. I.; Abell, C.; Korchev, Y. E.; Klenerman, D. J. Am. Chem. Soc. 2002, 124, 8810–8811
- 11. Piner, R. D.; Zhu, J.; Xu, F.; Hong, S.; Mirkin, C. A. Science 1999, 283, 661–663
- 12. Hong, S. H.; Mirkin, C. A. Science 2000, 288, 1808–1811
- 13. Hong, S. H.; Zhu, J.; Mirkin, C. A. Langmuir 1999, 15, 7897–7900
- 14. Hong, S. H.; Zhu, J.; Mirkin, C. A. Science 1999, 286, 523–525
- 15. Piner, R. P.; Hong, S.; Mirkin, C. A. Langmuir 1999, 15, 5457–5460
- 16. Ivanisevic, A.; Mirkin, C. A. J. Am. Chem. Soc. 2001, 123, 7887–7889
- 17. Wilson, D. L.; Martin, R.; Hong, S.; Cronin-Golomb, M.; Mirkin, C. A.; Kaplan, D.L. Proc. Natl. Acad. Sci. USA 2002, 98, 13660–13664
- 18. Demers, L. M.; Ginger, D. S.; Park, S.-J.; Li, Z.; Chung, S.-W.; Mirkin, C. A. Science 2002, 298, 1836–1838
- 19. Su, M.; Liu, X.; Li, S.-Y.; Dravid, V. P.; Mirkin, C. A. J. Am. Chem. Soc. 2002, 124, 1560–1561
- 20. Troughton, E. B.; Bain, C. D.; Whitesides, G. M.; Nuzzo, R. G.; Allara, D. L.; Porter, M. D. Langmuir 1988, 4, 365–385
- 21. Bain, C. D.; Whitesides, G. M. J. Am. Chem. Soc. 1989, 111, 7164–7175
- 22. Pique, A.; Chrisey, D. B., Eds. Direct–write technologies for rapid prototyping applications: sensors, electronics, and integrated power sources; Academic Press: San Diego, CA, 2002
- 23. Noy, A.; Miller, A. E.; Klare, J. E.; Weeks, B. L.; Woods, B. W.; DeYoreo, J. J. Nano Letters 2002, 2, 109–112
- 24. Lim, J. H.; Ginger, D. S.; Lee, K. B.; Heo, J.; Nam, J. M.; Mirkin, C. A. Angew. Chem. Int. Ed. Engl. 2003, 42, 2309–2312
- 25. Lee, K. B.; Lim, J. H.; Mirkin, C. A. J. Am. Chem. Soc. 2003, 125, 5588–5589
- 26. Lee, K.-B.; Park, S. J.; Mirkin, C. A. Science 2002, 295, 1702–1705
- 27. Vettiger, P.; Cross, G.; Despont, M.; Drechsler, U.; Durig, U.; Gotsmann, B.; Haberle, W.; Lantz, M. A.; Rothuizen, H. E.; Stutz, R.; Binnig, G. K. IEEE Transactions on Nanotechnology 2002, 1, 39–55
- 28. Chow, E. M.; Yaralioglu, G. G.; Quate, C. F.; Kenny, T. W. Appl. Phys. Lett. 2002, 80, 664–666
- 29. Sulchek, T.; Grow, R. J.; Yaralioglu, G. G.; Minne, S. C.; Quate, C. F.; Manalis, S.R.; Kiraz, A.; Aydine, A.; Atalar, A. Appl. Phys. Lett. 2001, 78, 1787–1789
- 30. Degertekin, F. L.; Hadimioglu, B.; Sulchek, T.; Quate, C. F. Appl. Phys. Lett. 2001, 78, 1628–1630
- 31. Zhang, M.; Bullen, D.; Chung, S.-W.; Hong, S.; Ryu, K. S.; Fan, Z.; Mirkin, C. A.; Liu, C. Nanotechnology 2002, 13, 212–217
- 32. Thorsen, T.; Maerkl, S. J.; Quake, S. R. Science 2002, 298, 580–584
- 33. Storho , J. J.; Mucic, R. C.; Mirkin, C. A. J. Cluster Sci. 1997, 8, 179–216
- 34. Storho , J. J.; Elghanian, R.; Mucic, R. C.; Mirkin, C. A.; Letsinger, R. L. J. Am. Chem. Soc. 1998, 120, 1959–1964
- 35. Niemeyer, C. M.; Burger, W.; Peplies, J. Angew. Chem. Int. Ed. 1998, 37, 2265– 2268
- 36. Niemeyer, C. M. Angew. Chem. Int. Ed. Engl. 2001, 40, 4128–4158
- 37. Park, S. J.; Taton, T. A.; Mirkin, C. A. Science 2002, 295, 1503–1506
- 38. Haes, A. J.; Van Duyne, R. P. J. Am. Chem. Soc. 2002, 124, 10596–10604
- 39. Taton, T. A.; Lu, G.; Mirkin, C. A. J. Am. Chem. Soc. 2001, 123, 5164–5165
- 40. Taton, T. A.; Mirkin, C. A.; Letsinger, R. L. Science 2000, 289, 1757
- 41. Parak, W. J.; Boudreau, R.; Le Gros, M.; Gerion, D.; Zanchet, D.; Micheel, C. M.; Williams, S. C.; Alivisatos, A. P.; Larabell, C. Adv. Mater. 2002, 14, 882–885
- 42. Demers, L. M.; Park, S. J.; Taton, T. A.; Li, Z.; Mirkin, C. A. Angew. Chem. Int. Ed. Engl. 2001, 40, 3071–3073
- 43. Adleman, L. M. Molecular computation of solutions to combinatorial problems. Science 1994, 266, 11 November
- 44. Lipton, R. J. DNA solution of hard computational problem. 1995, Science 268, 28 April
- 45. Ouyang, Q.; Kaplan, P. D.; Liu, S.; Libchaber, A. DNA solution of the maximal clique problem. Science 1997, 278, 17 October
- 46. Liu, Y., Xu, J. Pan, L., Wang, S. DNA Solution of a graph coloring problem. J. Chem. Inf. Comput. Sci. 2002, 42, 524–528
- 47. Smith L. M, Corn R. M, Condon A. E, Lagally M. G, Frutos A. G, Liu Q, Thiel A. J. A surface–based approach to DNA computation. J Comput Biol. 1998 Summer, 5(2):255–67
- 48. Frutos, A. G., Smith, L. M., and Corn, R. M. (1998) Enzymatic ligation reactions of DNA “words” on surfaces for DNA computing. J. Am. Chem. Soc. 120, 10277– 10282. York, Vol. 1
- 49. Liu, Q, Wang, L., Frutos, A. G., Condon, A.E. Corn, R. M. Smith, L. M. DNA computing on surfaces. Nature, Vol. 403, 13 Jan 2000 175–178
- 50. Sch¨oning, U. in Proc. 40th Ann. IEEE Conf. Found. Comp. Sci. (FOCS) 410–414 (IEEE Comp. Sci., Los Alamitos, California, 1999)
- 51. Ogihara M. Ray, A. DNA computing on a chip. Nature Vol 403, 143–144, 2000
- 52. Kinosita Jr, K., Yashida, R., Noji, H., Ishiwata, S., Yoshida, M. Cell 93, 21 (1998)
- 53. Noji, H., Yasuda, R., Yoshida, M., Kinosita Jr, K., Direct observation of the rotation of F1ATPase. Nature, 386, 299–302, 1997
- 54. Bachand, G.D. Montemagno, C. D. Constructing Organic/Inorganic NEMS Devices Powered by Biomolecular Motors. Biomedical Microdevices 2:3, 179–184, 2000
- 55. Yoshida M, Muneyuki E, Hisabori T. ATP synthase–a marvelous rotary engine of the cell. Nat Rev Mol Cell Biol. 2001 Sep;2(9):669–77
- 56. Noji H, Yasuda R, Yoshida M, Kinosita K Jr. Direct observation of the rotation of F1-ATPase. Nature, 1997, 386 :299–302.
- 57. Soong, R.K., Bachand, G.D., Neves, H.P., Olkhovets, A.G., Craighead, H.G., Montemagno, C.D. Powering a nanodevice with a biomolecular motor. Science, 290, 1555–, 2000
- 58. Bachand, G.D. Montemagno, C. D. Constructing Organic/Inorganic NEMS Devices Powered by Biomolecular Motors. Biomedical Microdevices 2:3, 179–184, 2000
- 59. Soong, R.K., Neves, H.P., Schmidt, J.J., Bachand, G.D., Montemagno, C.D.
- Engineering Issues in the fabrication of a hybrid nano–propeller system powered by F1–ATPase, Biomedical Microdevices, 3:1, 71–73, 2001118 Dan V. Nicolau et al.
- 60. Hess H, Vogel V. Molecular shuttles based on motor proteins: active transport in synthetic environments. J Biotechnol. 2001 Nov; 82(1):67–85
- 61. Yanagida, T., Nakase, M., Nishiyama, K. & Oosawa, F. (1984). Direct observation of motion of single F–actin filaments in the presence of myosin. Nature 307, 58–60
- 62. Spudich, J. A., Kron, S. J. & Sheetz, M. P. (1985) Nature, 315, 584–586
- 63.Suzuki, H., Oiwa, K., Yamada, A., Sakakibara, H., H. Nakayama and Mashiko
- S.1995. Linear arrangement of motor protein on a mechanically deposited fluoropolymer thin film. Jpn. J. Appl. Phys. 34:3937–3941
- 64. Suzuki, H., Yamada, A., Oiwa K., H. Nakayama and S. Mashiko. 1997. Control of actin moving trajectory by patterned poly(methylmethacrylate) tracks. Biophys.J.72:1997–2001
- 65. Nicolau, D.V., Suzuki, H., Mashiko, S., Taguchi, T., Yoshikawa, S. Movement of actin filaments on microlithographically–functionalized myosin tracks. Biophysical Journal, 77 (2), 99044–99065, 1999
- 66. Dennis, J. R., Howard, J. & Vogel, V. (1999). Molecular shuttles: directing the motion of microtubules on nanoscale kinesin tracks. Nanotechnology 10, 232–236
- 67. Mahanivong, C., Wright, J. P., Kekic, M., Pham, D. K., dos Remedios, C., Nicolau, D.V. Manipulation of the Motility of Protein Molecular Motors on Microfabricated Substrates. Biomedical Microdevices 4(2): 111–116; 2002
- 68. Clemmens, J.; Hess, H.; Howard, J.; Vogel, V. Analysis of Microtubule Guidance in Open Microfabricated Channels Coated with the Motor Protein Kinesin. Langmuir; 2003; 19(5); 1738–1744
- 69. Bunk, R., Klinth , J., Rosengren , J., Nicholls, I., Tagerud, S., Omling, P., Mansson, A., Montelius, L. Towards a ‘nano–tra c’ system powered by molecular motors. Microelectronic Engineering 67–68 (2003) 899–904
- 70. Turner, D. C., Chang, C., Fang K., S. L. Brandow and D. B. Murphy. 1995. Selective adhesion of functional microtubules to patterned silane surfaces. Biophys.J.69:2782–2789
- 71. Riveline, D., Ott, A., Julicher, F., Winkelmann, D. A., Cardoso, O., Lacapere, J.J., Magnusdottir, S., Viovy, J. L., Gorre-Talini, L. & Prost, J. (1998). Acting on actin: the electric motility assay. Eur Biophys J 27, 403–8
- 72. Hiratsuka, Y., Tada, T., Oiwa, K., Kanayama, T., Uyeda, T.Q. P. Controlling the Direction of Kinesin–Driven Microtubule Movements along Microlithographic Tracks. Biophysical Journal, 81 2001 1555–1561
- 73. Fulga, F., Myhra, S., Nicolau, Jr. D. V., D.V. Nicolau, Interrogation of the dynamics of magnetic microbeads on the meso–scale via electromagnetic detection. Smart Materials & Structures, 11 (5) 722–727, 2002
- 74. Hess, H., Clemmens, J., Qin, D., Howard, J. & Vogel, V. (2001). Light–Controlled Molecular Shuttles Made from Motor Proteins Carrying Cargo on Engineered Surfaces. Nano Letters 1, 235–239
- 75. Hess, H.; Howard, J.; Vogel, V. A Piconewton Forcemeter Assembled from Microtubules and Kinesins. Nano Lett. 2002; 2(10); 1113–1115
- 76. Hess, H.; Clemmens, J.; Howard, J.; Vogel, V. Surface Imaging by Self–Propelled Nanoscale Probes. Nano Lett. 2002; 2(2); 113–116
- 77. Nicolau, D. V., Jr.; Nicolau, D. V. Computing with the Actin–Myosin molecular motor system. In Biomedical Applications of Micro– and Nanoengineering. Nicolau, Dan V. (Ed.) SPIE Proc. 4937, 219–225, 2002
Есть ли ДНК чипы в вакцинах от COVID-19?
Пока нет. В вакцинах на сегодняшний день есть многочисленные oligonucleotide microarrays, которые предназначены для управления на уровне РНК, Long terminal repeats промежуточных соединений РНК, встроенных через вакцину полиомиелита эндовирусов SV40. Эти microarrays работают в течение длительного времени (на длительном временном интервале).
Связующими элементами управления вакцин являются ретротранспозоны. Ретротранспозоны (называемые транспозонами класса I или транспозонами через промежуточные соединения РНК ) представляют собой тип генетических компонентов, которые копируют и вставляют себя в разные места генома (транспозон ) путем преобразования РНК обратно в ДНК посредством процесса обратной транскрипции с использованием промежуточных продуктов транспозиции РНК.
Посредством обратной транскрипции ретротранспозоны быстро амплифицируются и становятся изобильными в геномах эукариот, таких как кукуруза (49–78%) и Homo sapiens (42%). Они присутствуют только у эукариот, но имеют общие черты с ретровирусами , такими как ВИЧ.
В 2021 году нам не будут вводить микрочипы через иглу вакцинного шприца.
Набор своих microarrays мы получим через элементы генно модифицированного питания. Эта первая фаза будет состоять из модифицированных генов, сои, рапса, картофеля, томатов, лимонной кислоты, сахарного песка, говядины (коров кормят генно модифицированным рапсом), свинины (ученые уже создали свиней, которые могут стать первыми генно-модифицированными животными, одобренными для употребления в пищу. От экспериментальных аналогов он отличается тем, что использует простую правку генов, а не смешение генетических материалов разных организмов.) Рапс — растение из рода капусты, гибрид огородной капусты и травы сурепки. Семена рапса содержат много масла, но оно неприятно на вкус и применяется главным образом для технических нужд, а также как добавка в корм скоту и в самые дешевые маргарины.

А вот вторая фаза будет включать в себя различные системы идентификации и аутентификации каждого индивидуума, должна закончиться на разных территориях в 2025 -2030 году может включать в себя наносистемы, способные работать в режиме физического ансамбля корреляционных приемников и, получая электромагнитную энергию от плотных электромагнитных полей 5G систем связи и запускать моторы.
И к этому времени уже будут готовы алгоритмы Earth Global Computer от Microsoft и искусственный интеллект удаленного распознавания. Вызывают сомнения только 2 вот этих коррелирующих картинки


Динамика графика смертности в США от SARS-COV-2 подозрительно напоминает график в модели случайного блуждания в беспроводной сети 5G с интерференцией… А сеть 5G в больших городах Соединенных штатов Америки уже развернута и функционирует. К чему бы это?
ID2020.org, Global Alliance for Vaccines and Immunization, распознавание иммунных паспортов мобильных элементов беспроводной сети 5G методом мгновенного иммуно ферментного анализа антител к двухцепочечным ДНК (dsDNA) путем сбора и анализа ответов статистического ансамбля nanoarrays — классическая пеленгация на базе 5G — аналог метки радиочастотной идентификации на базе эндогенного SARS-COV-2 — это Mortal Combat 22 века?
Информационные потоки снова замкнулись. Только вот есть одна маленькая проблема. C фактором SEIR q=4 для Preductor-а типа SARS-COV-2 в первые 17 дней распространения свой штамм вируса получил почти каждый житель планеты Земля. Просто дозы смертельного патогена и уровень вирусной нагрузки сильно различались. А дальше заработали эволюционные законы, принцип защиты Ле-Шателье и брюсселятор сложной распределенной биофизической системы….
Чем будет сильнее иммунный ответ организма на коронавирус, тем будет более неблагоприятным прогноз. Биосфера планеты это тоже организм, так что чем сильнее ответ коронавирусу, тем масштабнее аутоиммунный ответ по принципу Ле-Шателье…
Так что богатые тоже плачут… Боже, храни королеву…